
The Amino Acids
Notes from the edx/harvardX biochemistry class (2015)
A protein is a linear polymer, or polypeptide, of amino acids, each linked by a peptide bond. Proteins are responsible for the many of the essential functions of life.
There are 22 proteinogenic amino acids. These amino acids have four different groups attached to its central carbon, known as the α-Carbon: an amino group, a carboxyl group, and a side chain R (variable).

At the physiological pH of ~7.4, the amino group and carboxyl groups are ionized (AG+, CG–), making the amino acid neutral and thus zwitterionic: a molecule with both positively/negatively charged components.

An amino acid is also chiral because it has four different groups off the α-carbon. Chiral molecules are considered optically active—that is, they rotate plane-polarized light.

In terms of chirality, the amino acids used in nature are l-amino acids, as opposed to d-amino acids. Note that our bodies cannot process d-amino acids!

L-amino acids when viewed head-on have their N-terminus on the left and C-terminus on the right. The α-carbon will be on top with side chains pointing toward you.

Recall the chirality of carvone from the intro lecture, that the enantiomers S-(+) and R-(–) Carvone have different tastes: one is spicy (caraway), the other sweet (spearmint).
The amino acids can be classified in many different ways. The most common grouping is shown below. Note the each aminos acid is identified by a single letter, the sequence of which is used to identify a given protein. For example, …THVLPFEKINEGFDLLR…

Polarity is an important attribute because it affects the types of non-covalent interactions that can be formed. Relevant to proteins are the following three:
- van der Waals: weakest, acts on short distances, present between any two atoms in close proximity. The distance at which the energy is minimal is the van der Waal radius. Allows geckos to cling to surfaces vertically, btw.
- Salt Bridges: between pairs of charged ions. Strongest in terms of binding energy: 10x > van der Waals, depending on the polarity of the environment. Salt Bridges are a combination of ionic bonds and hydrogen bonds.
- Hydrogen Bonds: 2x-5x > van der Waals. Only occurs between polar groups and permanent dipole (one a H-bond donor, one an acceptor). Unique in that they are directional, unlike others.
Two examples that show the different functionalities of AA side chains:
- Aspartate (Asp) vs. Glutamate (Glu). Asp and Glu only differ by one methylene in the linear section. Glu is longer, which gives Glu many more conformations (“rotamers”) and thus affords it more fitting opportunities. But this comes at entropic cost: once it bonds to a ligand it will have lost more conformational entropy than Aspartate. If you don’t know what “bonds to a ligand” means yet, it’ll be explained in later sections. As it pertains to entropy, it just means “once the thing is forced to hold still.”

- Histidine (His/H) is often found in the active sites of enzymes. Histidine can exist in the multiple forms, depending on the solution pH, from acidic to basic. With a pKa ~6.0, it can be either charged or neutral.

When H is in its apolar conformation, it has two configurations around the imidazole group.

The ability to be both polar and apolar makes it relatively easy to move protons on and off the side chain, which means it can live inside the hydrophobic protein core or on the surface surrounded by solvent. It’s also ideal for active sites.
Here are pKa‘s for a few other amino acids:

Dr. Gaudet’s Food for Thought: why would Tyrosine be so much more acidic than Serine, even though they’re both alcohols?
The thinking: Tyrosine has a resonance cyclic structure and it’s aromatic, which makes for an extremely stable molecule. Its conjugate base (which forms a phenol group) is therefore highly stable, much more so that the corresponding conjugate base on Serine. The more stable the base, the stronger the acid, thus Tyrosine wins the acid war here. More on this here. And here especially.

Now, most amino acid side chains are formed of carbon, nitrogen, oxygen, and hydrogen, but two also contain sulfur: Methionine, a hydrophobic residue, and cysteine.

Cysteine is quite reactive under physiological conditions and can easily be oxidized to form sulfide bonds. These conditions are usually found on the extracellular side.
Back to chirality; note that all amino acids are chiral except two:
- Glycine, which has R = H. So it’s not chiral (3 not 4 groups). The smaller R gives it fewer conformational restrictions, important to protein folding.
- Proline, a cyclic AA whose R is covalently linked to the amino group and thus, more conformational restricted.
Amino acid classifications can be shown in a Venn diagram.
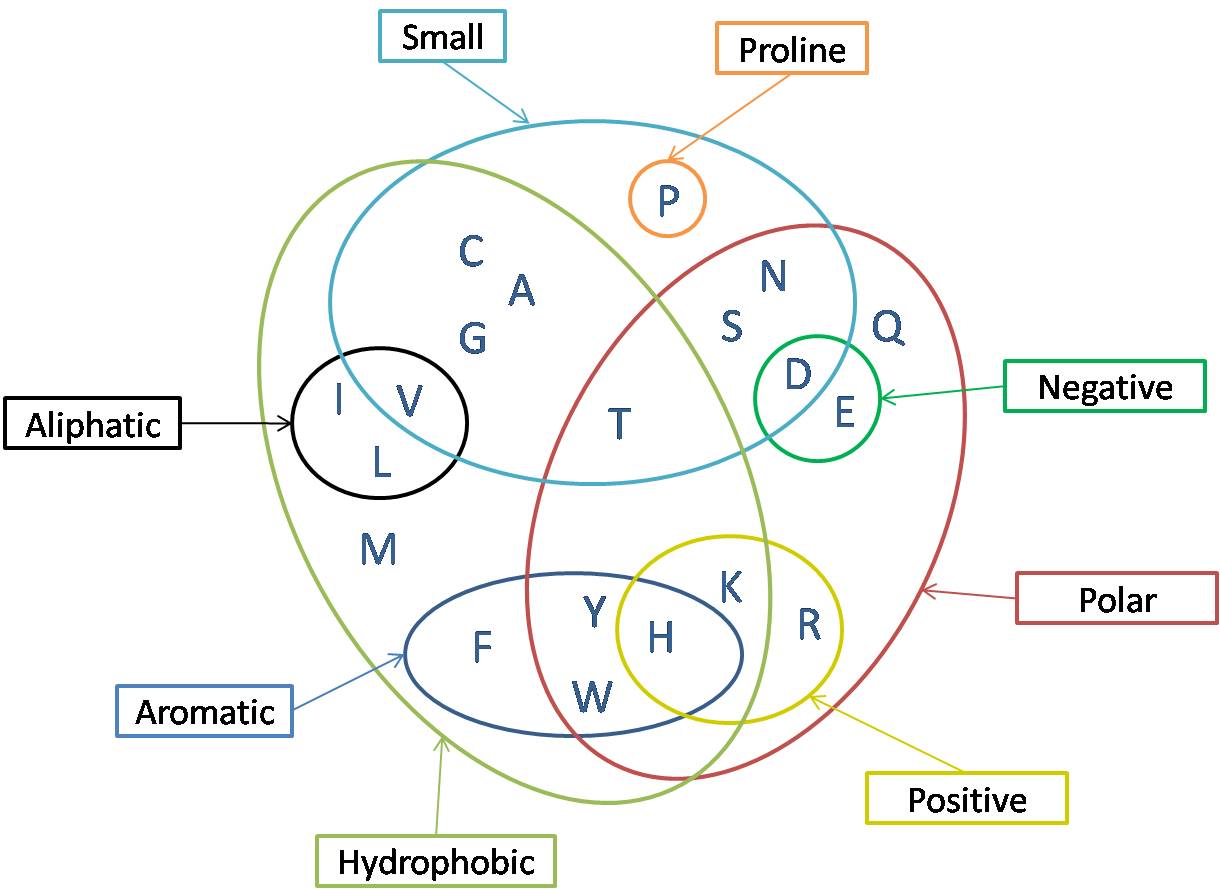
Aliphatic: Alanine (A), Valine (V), Methionine (M), Leucine (L), Isoleucine (I)
Aromatic: Tryptophan (W), Phenylalanine (F), Tyrosine (Y)
Polar: Threonine (T), Serine (S), Asparagine (N), Glutamine (Q), Y, and C
Charged, Basic: Histidine (H), Arginine (R), Lysine (K)
Charged, Acidic: Aspartic Acid (D), Glutamic Acid (E)
Conformationally unique: Proline (P), Glycine (G)
Chemically unique: Cysteine (C), Histidine (H)
Apolar (hydrophobic, often buried in the core): A, I, L, F, V, P, G
Essential Amino Acids: Phenylalanine, Valine, Threonine, Tryptophan, Isoleucine, Methionine, Histidine, Arginine, Leucine, and Lysine (Pvt. Tim Hall)
Nonessential Amino Acids: Alanine, Asparagine, Aspartic Acid, Glutamic Acid.
Conditional Amino Acids: Arginine (essential in children), Cysteine, Glutamine, Glycine, Proline, Serine, and Tyrosine.
Interesting example of crossover is Lysine. It is charged at 7.4 because its R contains a positive charge. It’s polar because it can form hydrogen bonds. Yet, it can also be non-polar since the rest of the R is aliphatic, or non-polar. So in some situations, it will behave as hydrophobic.

Here’s a tremendous resource on the lore of Amino Acids: http://www.biochem.umd.edu/biochem/kahn/teach_res/amino_acids/


You May Also Like

Protein Classifications

The Ten Steps of Glycolysis
